
 Title: The Greatest Show on Earth: The Evidence for Evolution
Title: The Greatest Show on Earth: The Evidence for Evolution
Author(s): Richard Dawkins
Release year: 2009
Publisher: Transworld
Why in Database: Very turtle position! We had to make a certain selection of texts, the publisher might consider throwing 8-9 pages of text as a kind of piracy. So we decided to throw in four normal fragments that mention turtles, two fragmenrs from the nine-page section mentioned, two fragments of colored inserts and two footnotes.
The first turtle element tells about the Galapagos, how they got their name:
By the way, the bird itself is named after the islands,* not the other way around as with the Galapagos Islands, whose name comes from a Spanish word for tortoise.
The second fragment addresses the problem with the term “reptiles”, turtles are of course mentioned in this context:
Cladistically inclined zoologists avoid the word ‘reptiles’ altogether, splitting them into Archosaurs (crocodiles, dinosaurs and birds), Lepidosaurs (snakes, lizards and the rare Sphenodon of New Zealand) and Testudines (turtles and tortoises). Non-cladistically inclined zoologists are happy to use a word like ‘reptile’ because they find it descriptively useful, even if it does artificially exclude the birds.
Another mention is a turtle in the list of various animals that returned from land to water:
Other animals that have returned from land to water, at least some of the time, are pond snails, water spiders, water beetles, crocodiles, otters, sea snakes, water shrews, Galapagos flightless cormorants, Galapagos marine iguanas, yapoks (aquatic marsupials from South America), platypuses, penguins and turtles.
The next two long fragments comes from the aforementioned huge fragment about turtles:
I now want to turn to another group of animals that returned from the land to the water: a particularly intriguing example because some of them later reversed the process and returned to the land a second time! Sea turtles are, in one important respect, less fully given back to the water than whales or dugongs, for they still lay their eggs on beaches. Like all vertebrate returners to the water, turtles haven’t given up breathing air, but in this department some of them go one better than whales. These turtles extract additional oxygen from the water through a pair of chambers at their rear end that are richly supplied with blood vessels. One Australian river turtle, indeed, gets the majority of its oxygen by breathing (as an Australian would not hesitate to say) through its arse.
Before going any further, I can’t escape a tiresome point of terminology, and a regrettable vindication of George Bernard Shaw’s observation that ‘England and America are two countries divided by a common language.’ In Britain, turtles live in the sea, tortoises live on land and terrapins live in fresh or brackish water. In America all these animals are ‘turtles’, whether they live on land or in water. ‘Land turtle’ sounds odd to me, but not to an American, for whom tortoises are the subset of turtles that live on land.
Some Americans use ‘tortoise’ in a strict taxonomic sense to refer to the Testudinidae, which is the scientific name for modern land tortoises. In Britain, we’d be inclined to call any land-dwelling chelonian a tortoise, whether it is a member of the Testudinidae or not (as we shall see, there are fossil ‘tortoises’ that lived on land but are not members of the Testudinidae). In what follows, I’ll try to avoid confusion, making allowance for readers in Britain and America (and Australia, where the usage is different again), but it’s hard. The terminology is a mess, to put it mildly. Zoologists use ‘chelonians’ for all these animals, turtles, tortoises and terrapins, whichever version of English we speak.
The most instantly noticeable feature of chelonians is their shell. How did it evolve, and what did the intermediates look like? Where are the missing links? What (a creationist zealot might ask) is the use of half a shell? Well, amazingly, a new fossil has just been described, which eloquently answers that question. It made its debut in the journal Nature in the nick of time before I had to hand this book over to the publishers. It was an aquatic turtle, found in late Triassic sediments in China, and its age is estimated at 220 million years. Its name is Odontochelys semitestacea, from which you may deduce that, unlike a modern turtle or tortoise, it had teeth, and it did indeed have half a shell. It also had a much longer tail than a modern turtle or tortoise. All three of these features mark it out as prime ‘missing link’ material.
The belly was covered by a shell, the so-called plastron, in pretty much the same way as that of a modern sea turtle. But it almost completely lacked the dorsal portion of the shell, known as the carapace. Its back was presumably soft, like a lizard’s, although there were some hard, bony bits along the middle above the backbone, as in a crocodile, and the ribs were flattened, as though ‘trying’ to form the evolutionary beginnings of a carapace.
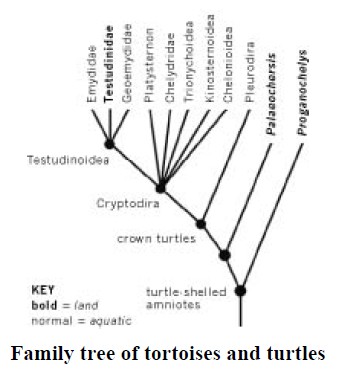
If you draw out the family tree of all modern turtles and tortoises, based on molecular and other comparisons, nearly all the branches are aquatic (normal type). Land tortoises are represented by bold type, and you can see that today’s land tortoises constitute a single branch, the Testudinidae, deeply nested within rich branchings of otherwise aquatic chelonians. All their close cousins are aquatic.
Modern land tortoises are a single twig on the bush of otherwise aquatic turtles. Their aquatic ancestors turned turtle and trooped back on to the land. This fact is compatible with the hypothesis that the shell evolved in water, in a creature like Odontochelys. But now we have another difficulty. If you look at the family tree, you’ll notice that, in addition to the Testudinidae (all modern land tortoises) there are two fossil genera of fully shelled animals called Proganochelys* and Palaeochersis. These are drawn as landdwellers, for reasons we shall come to in the next paragraph. They lie right outside the branches representing the water turtles. It would seem that these two genera are anciently terrestrial.
Before Odontochelys was discovered, these two fossils were the oldest known chelonians. Like Odontochelys they lived in the late Triassic, but about 15 million years later than Odontochelys. Some authorities have reconstructed them as living in fresh water, but recent evidence does indeed place them on land, as indicated by bold type on the diagram. You might wonder how we tell whether fossil animals, especially if only fragments are found, lived on land or in water. Sometimes it’s pretty obvious.
Ichthyosaurs were reptilian contemporaries of the dinosaurs, with fins and streamlined bodies. The fossils look like dolphins and they surely lived like dolphins, in the water. With turtles and tortoises it is a little less obvious. As you might expect, the biggest giveaway is their limbs. Paddles really are rather different from walking legs. Walter Joyce and Jacques Gauthier, of Yale University, took this commonsense intuition and provided the numbers to support it. They took three key measurements in the arm and hand bones of seventy-one species of living chelonians. I’ll resist the temptation to explain their elegant calculations, but their conclusion was clear. These animals had had walking legs, not paddles. In British English, they were ‘tortoises’, not ‘turtles’. They lived on land. They were only distant cousins, however, of modern land tortoises.
Now we seem to have a problem. If, as the authors of the paper describing Odontochelys believe, their half-shelled fossil shows that the shell evolved in water, how do we explain two genera of fully shelled ‘tortoises’ on land, 15 million years later? Until the discovery of Odontochelys, I would not have hesitated to say that Proganochelys and Palaeochersis were representative of the land-dwelling ancestral type before the return to water. The shell evolved on land. Some shelled tortoises returned to the sea, as seals, whales and dugongs were later to do. Others stayed on land, but went extinct. And then some sea turtles returned to the land, to give rise to all modern land tortoises. That’s what I would have said – indeed what I did say in the earlier draft of this chapter that preceded the announcement of Odontochelys.
But Odontochelys throws speculation back into the melting pot. We now have three possibilities, all equally intriguing.
1 Proganochelys and Palaeochersis might be survivors of the land-dwelling animals that had earlier sent some representatives to sea, including the ancestors of Odontochelys. This hypothesis would suggest that the shell evolved on land early, and Odontochelys lost the carapace in the water, retaining the ventral plastron.
2 The shell might have evolved in water, as the Chinese authors suggest, with the plastron over the belly evolving first, and the carapace over the back evolving later. In this case, what do we make of Proganochelys and Palaeochersis, who lived on land after Odontochelys lived, with its half shell, in water? Proganochelys and Palaeochersis might have evolved the shell independently. But there is another possibility:
3 Proganochelys and Palaeochersis might represent an earlier return from the water to the land. Isn’t that a startlingly exciting thought? We are already pretty confident of the remarkable fact that the turtles accomplished an evolutionary doubling back to the land: an early marque of land ‘tortoises’ went back to the watery environment of their even earlier fish ancestors, became sea turtles, then returned to the land yet again, as a new incarnation of land tortoises, the Testudinidae. That we know, or are nearly certain of. But now we are facing up to the additional suggestion that this doubling back happened twice! Not just to spawn the modern tortoises, but much longer ago, to give rise to Proganochelys and Palaeochersis in the Triassic.
In another book I described DNA as ‘the Genetic Book of the Dead’. Because of the way natural selection works, there is a sense in which the DNA of an animal is a textual description of the worlds in which its ancestors were naturally selected. For a fish, the genetic book of the dead describes ancestral seas. For us and most mammals, the early chapters of the book are all set in the sea and the later ones all out on land. For whales, dugongs, marine iguanas, penguins, seals, sea lions and turtles, there is a third section of the book which recounts their epic return to the proving grounds of their remote past, the sea.
But for the land tortoises, perhaps twice independently on two widely separated occasions, there is yet a fourth section of the book devoted to a final – or is it? – re-emergence, yet again to the land. Can there be another animal for which the genetic book of the dead is such a palimpsest of multiple evolutionary Uturns?
As a parting shot, I cannot help wondering about those freshwater and brackish water forms (‘terrapins’), which are close cousins of the land tortoises. Did their ancestors move directly from the sea into brackish and then fresh water? Do they represent an intermediate stage on the way from the sea back to the land? Or is it possible that they constitute yet another doubling-back to the water from ancestors that were modern land tortoises? Have the chelonians been shuttling back and forth in evolutionary time between water and land? Could the palimpsest be even more densely over-written than I have so far suggested?
The next fragment is a little bit about Darwin and the Galapagos, but mostly about the turtle diversity on these islands:
So it is, all over the world. The fauna and flora of a particular region are just what we should expect if, to quote Darwin on the finches that now bear his name, ‘one species had been taken and modified for different ends’.
The Vice-Governor of the Galapagos Islands, Mr Lawson, intrigued Darwin by informing him that the tortoises differed from the different islands, and that he himself could with certainty tell from which island any one was brought. I did not for some time pay sufficient attention to this statement, and I had already partially mingled together the collections from two of the islands. I never dreamed that islands, about fifty or sixty miles apart, and most of them in sight of each other, formed of precisely the same rocks, placed under a quite similar climate, rising to a nearly equal height, would have been differently tenanted.
All the Galapagos giant tortoises are similar to a particular mainland species of land tortoise, Geochelone chilensis, which is smaller than any of them. At some point during the few million years that the islands have existed, one or a few of these mainland tortoises inadvertently fell in the sea and floated across.
How could they have survived the long and doubtless arduous crossing? Surely most of them didn’t. But it would have only taken one female to do the trick. And tortoises are astonishingly well-equipped to survive the crossing.
The early whalers took thousands of giant tortoises from the Galapagos islands away in their ships for food. To keep the meat fresh, the tortoises were not killed until needed, but they were not fed or watered while waiting to be butchered. They were simply turned on their backs, sometimes stacked several deep, so they couldn’t walk away. I tell the story not in order to horrify (although I have to say that such barbaric cruelty does horrify me), but to make a point. Tortoises can survive for weeks without food or fresh water, easily long enough to float in the Humboldt Current from South America to the Galapagos archipelago. And tortoises do float.
Having reached and multiplied upon their first Galapagos island, the tortoises would with comparative ease – again accidentally – have island-hopped the much shorter distances to the rest of the archipelago by the same means. And they did what many animals do when they arrive on an island: they evolved to become larger. This is the long-noticed phenomenon of island gigantism (confusingly, there is an equally well-known phenomenon of island dwarfism).*
If the tortoises had followed the pattern of Darwin’s famous finches, they would have evolved a different species on each of the islands. Then, after subsequent accidental driftings from island to island, they would have been unable to interbreed (that’s the definition, remember, of a separate species) and would have been free to evolve a different way of life uncontaminated by genetic swamping.
You could say that the different species’ incompatible mating habits and preferences constitute a kind of genetic substitute for the geographic isolation of separate islands. Though they overlap geographically, they are now isolated on separate ‘islands’ of mating exclusivity. So they can diverge yet further. The Large, the Medium and the Small Ground Finch originally diverged on different islands; the three species now coexist on most of the Galapagos islands, never interbreeding and each specializing in a different kind of seed diet.
The tortoises did something similar, evolving distinctive shell shapes on the different islands. The species on the larger islands have high domes. Those on the smaller islands have saddle-shaped shells with a high-lipped window at the front for the head. The reason for this seems to be that the large islands are wet enough to grow grass, and the tortoises there are grazers. The smaller islands are mostly too dry for grass, and the tortoises resort to browsing on cactuses. The high-lipped saddle shell allows the neck to reach up to the cactuses which, for their part, grow higher in an evolutionary arms race against the browsing tortoises.
The tortoise story adds to the finch model the further complication that, for tortoises, volcanoes are islands within islands. Volcanoes provide high, cool, damp, green oases, surrounded at low altitude by dry lava fields which, for a grazing giant tortoise, constitute hostile deserts. Each of the smaller islands has a single large volcano and its own single species (or sub-species) of giant tortoise (except those few islands that have none at all). The big island of Isabela (‘Albemarle’ to Darwin) consists of a string of five major volcanoes, and each volcano has its own species (or sub-species) of tortoise. Truly, Isabela is an archipelago within an archipelago: a system of islands within an island. And the principle of islands in the literal geographical sense, setting the stage for the evolution of islands in the metaphorical genetic sense of species, has never been more elegantly demonstrated than here in the archipelago of Darwin’s blest youth.*
The next two mentions appear in the footnotes (which are in this long-mentioned passage):
The rule seems to be that, on islands, big animals get smaller (for example, there were dwarf elephants the height of a large dog on Mediterranean islands such as Sicily and Crete) while small animals get bigger (as in the Galapagos tortoises). There are several theories for this divergent tendency, but the details would take us too far afield.
* These paragraphs, on giant tortoises, are extracted from an article that I wrote on a boat called the Beagle (not the real one, which is unfortunately long extinct) in the Galapagos archipelago, and published in the Guardian on 19 February 2005.
The next two pieces come from colored inserts with illustrations – both illustrations (one double) are also in the gallery below:
Some groups – as suggested by the fascinating Odontochelys semitestacea, the proto-turtle without a top shell (f) – having returned to the water, may even have made another return to the land later.
The giant tortoises of Galapagos vary from island to island. The saddleback shape (e), unlike the dome shape of grazing tortoises (f), is characteristic of islands where the tortoises browse on cactuses and therefore have to stretch their necks high.




Author: XYuriTT